
土壤墒情对小叶扶芳藤的光合作用中的光响应的影响
土壤墒情和光照强度是影响植被生长及分布的重要生态因子,两者对植物的光合、蒸腾及有机物运输等生理生态过程产生作用,从而影响植物对水资源和光能的利用效率。随着大气臭氧层的减少,光因子作为环境胁迫的作用越来越突出。同时干旱缺水是制约植被恢复与农林业生产最为关键的生态因子,随着水资源危机和干旱化危害的不断加剧,节水农林业生产模式的发展日益被重视。植物如何适应全球气候变化所引起的土壤干旱和光因子胁迫是人们关注的问题之一.近年来对光合生产的研究主要集中在农作物和经济林领域,光合特性及其生态影响因子的时空动态成为研究的主要内容之一,水分胁迫下其气体交换特性的研究多涉及到三、四个水分梯度,对多级水分胁迫梯度下的光合生理指标变化规律研究较少。
藤本植物小叶扶芳藤为常绿耐寒树种,适应性强,攀附能力强、可塑性好,在我国各地均有栽培,是应用于城市立体绿化、园林造景、水土保持等方面良好的植物材料.目前有关小叶扶芳藤的研究,多集中在园林绿化及栽培技术等方面,有关生理生态学特性方面的研究还处于起步阶段。本研究主要探讨多级土壤墒情梯度下小叶扶芳藤苗木生理生态过程及其对光强的响应,阐明小叶扶芳藤光合作用、蒸腾作用、水分利用效率、光能利用效率对土壤干旱与强光胁迫的响应规律,确定维持较高光合速率和水分利用效率的适宜土壤水分及光照强度范围,为小叶扶芳藤在植被恢复与重建中的应用提供理论依据。
1 材料和方法
1.1 试验地概况
研究区位于山西省中阳县的车鸣峪林场,属黄河流域典型的黄土丘陵沟壑区,地处东经111°04′~111°21′、北纬37°03′~37°20′.该区属暖温带亚干旱区大陆性明显的季风气候,气候干燥.多年平均降水量为500~550 mm,多集中在7~9月份(占全年降水量的70%以上),多年平均蒸发量为1 019.7mm.多年平均气温6℃左右,极端最高气温35.6℃(1994年6月16日),极端最低气温―24.3℃(1980年1月30日),无霜期125~150 d,≥10℃年积温为2 500~3 000℃.其土壤以褐土较多,其次为棕壤,且发育差,水土流失严重.它属于森林草原灌丛植被区,现有树种少,林地大部分是疏林地,林分稳定性差.
1.2 试验材料与水分处理设计
选用3年生小叶扶芳藤苗木,布设6个重复(布设6盆),盆内土壤为褐土,盆钵直径40 cm,深50cm.2006年3月份进行苗木盆栽,7月中旬进行实验观测.用环刀法测得盆栽试验土壤的田间持水量为25.3%,土壤容重为1.2 g/cm3.采用田间环境条件下人为给水和自然耗水相结合的方法,获取盆栽土壤不同的水分梯度.具体做法是:选取生长健壮较一致的小叶扶芳藤盆栽苗3株(每盆1株),试验观测2 d前(7月14日)浇水,使土壤墒情饱和,利用土壤墒情速测仪监测土壤墒情的变化.2 d后获得初期水分梯度,重量含水量(Masswater content,MWC:%)为21.4%,相对含水量(Relative water content,RWC:%)为84.8%,进行第1次生理参数的测定.以后通过自然耗水,每隔1~2 d获取1个水分梯度,共7个梯度(表1),分别进行生理参数光响应过程测定。
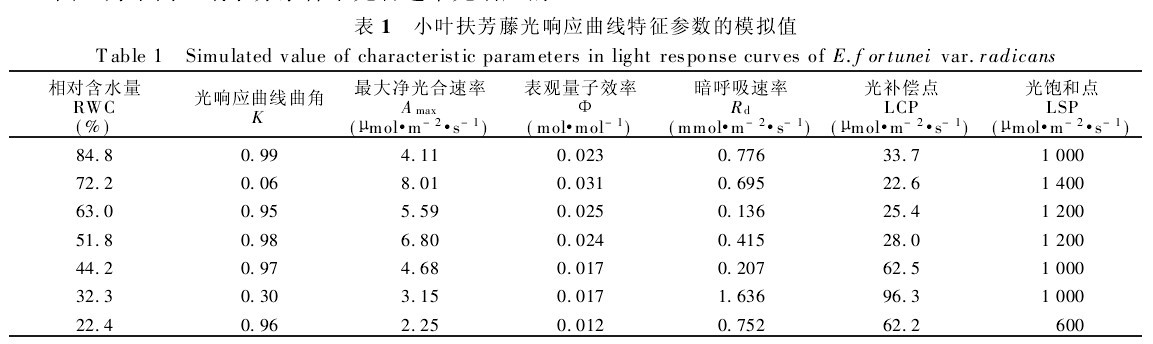
表1 小叶扶芳藤光响应曲线特征参数的模拟值
1.3 光响应过程测定
从每盆试验植株中部选6片生长健壮的成熟叶片,应用英国PPS公司生产的CIRAS-2型光合作用系统测定不同土壤水分下生理参数的光响应过程,为了尽量减少外界光照波动所造成的影响,测定时间选择在完全晴朗天气下的9:00~10:00左右时进行,每个叶片重复3次,取平均值.测定时,使用大气CO2浓度,利用人工光源将光合有效辐射强度控制在1 800、1 600、1 400、1 200、1 000、800、600、400、300、200、150、100、50、20μmol/m-2s-1等共14个梯度.每个梯度下控制测定时间为180 s.仪器自动记录净光合速率(Pn:μmol!m-2!s-1)、光合有效辐射(PAR:μmol!m-2!s-1)、蒸腾速率(Tr:mmol!m-2!s-1)等生理参数.叶片水分利用效率(WUE:μmol!m-2!s-1)、叶片瞬时光能利用效率(LUE)分别用公式计算,即:WUE=Pn/Tr,LUE=Pn/PAR.
式中的相对含水量(RWC)为重量含水量与田间持水量的比值.
1.4 计算及分析方法
绘制光合作用的光响应曲线(Pn-PAR曲线),采用非直角双曲线模型进行模拟:
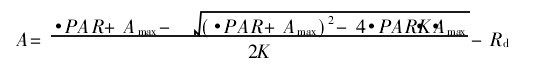
非直角双曲线模型进行模拟
式中,A为净光合速率,PAR为光合有效辐射,Amax为最大净光合速率,Ф、K、Rd分别为表观量子效率、光响应曲线曲角、暗呼吸速率.对Pn-PAR曲线的初始部分(PAR<200μmol!m-2!s-1)进行线性回归,可求得暗呼吸速率(Rd:mmol!m-2!s-1)、表观量子效率(Ф:mol!mol-1)、光补偿点( LCP:μmol/m-2s-1)等参数,光饱和点(LSP:μmol!m-2!s-1)根据光响应曲线实测数据点的走势估计(表1).利用统计分析软件Spss、Excel分析光响应模拟过程,做聚类分析及方差分析.
2 结果与分析
2.1 不同土壤水分条件下光响应曲线特征参数
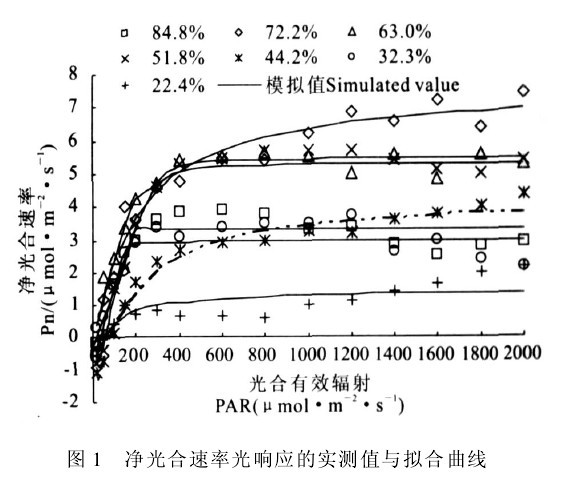
图1 净光合速率光响应的实测值与拟合曲线
图1为不同土壤墒情条件下光合速率光响应的模拟值与实测值,可以看出模拟值与实测值的变化趋势基本一致,模拟方程的决定系数R2均在0.95以上,说明模型可以较准确地模拟叶片光合速率的光响应规律,但从实际拟合效果看,Amax模拟值稍大于实测值,可能与该类型曲线特点及小叶扶芳藤超过光饱和点后,在高光强下Pn下降缓慢有关.
曲角K是表示光响应曲线形状的参数,从表1和图1可以看出,在土壤RWC为72.2%、32.3%时,K值较小,没有出现光饱和现象,而在其它土壤水分条件下K值变化不大且维持在较高值接近于1,呈现出达到光饱和状态的特征,说明小叶扶芳藤的光响应曲线达到最大值加快,在光饱和点以前几乎呈一条上升的直线,净光合通量增加较快,在光饱和点以后为光合速率维持在较高值且变化不大的直线。
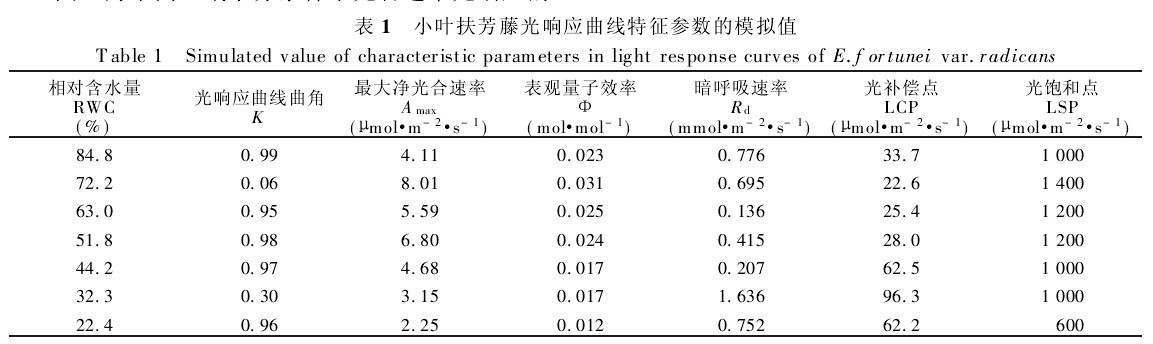
表1 小叶扶芳藤光响应曲线特征参数的模拟值
由表1可见,不同水分条件下小叶扶芳藤光响应曲线的特征参数存在着差异.随着土壤湿度的增高,Φ也随之增高,在RWC为72.2%时,Amax、Φ均达到最高值,土壤水分过多时(RWC为84.8%),Amax、Φ下降较大.在不同水分条件下,Φ值都小于理论上的最大量子效率(在0.08~0.125mol!mol-1)(表1).
而Rd则变化规律复杂,各土壤湿度下对光合产物的消耗表现出较大的差异,在土壤湿度较大(RWC为84.8%)及重度干旱胁迫下(RWC为32.3%),光合产物的消耗较大:而在轻度干旱胁迫下(RWC为63.0%),Rd较小,可减少呼吸作用对光合产物的消耗,有利于干物质的积累,为其提高生物产量奠定了良好的物质基础。
在最适环境条件下,叶片的最大光合速率表示了叶片的最大光合能力,表观量子利用效率反映了叶片对光能的利用情况,尤其是对弱光的利用能力,呼吸速率则与叶片的生理活性有关。通过对不同水分条件下小叶扶芳藤光响应曲线特征参数分析,表明在土壤RWC为72.2%时光合同化潜力最大,对弱光的利用能力强,生理活性高,在弱光条件下,仍维持较高的光合能力。
小叶扶芳藤LCP在22.5~96.6μmol/m-2s-1之间变动,介于阴性植物(LCP<20μmol!m-2!s-1)与阳性植物(50μmol!m-2!s-1
2.2 不同水分条件下净光合速率光响应
图1表明,在各土壤湿度下,Pn对PAR的响应表现为,一定光强范围内,Pn随着PAR的增强而增大,当光强超过一定范围后,这种增大的趋势逐渐减弱.具体表现:在低光强下(PAR<400μmol/m-2s-1),Pn对PAR变化反应最敏感,Pn随着PAR的增强急剧上升.此时在同一光强下,各土壤水分下Pn值变化幅度也较大,如PAR为200μmol/m-2s-1时,Pn变幅在0.73(RWC为22.4%)~4.23(RWC为63.0%)μmol/m-2s-1.当RWC为44.2%和72.2%时,Pn随着PAR的增强而上升,一直没有出现光饱和现象.在其它水分条件下,随着PAR的持续增强,Pn达到最大值,即出现光饱和点(LSP),超过LSP后,Pn有所下降,但仍能保持较高值,此时PAR在600~1 600μmol!m-2!s-1左右.Pn对RWC的响应表现为,在RWC<72.2%时,Pn随着RWC的增大而上升:此后随着RWC的增大,Pn反而下降,因此RWC为72.2%可作为Pn变化的转折点。
分析表明,如果在强光下提高小叶扶芳藤的光合生产力,必须有适宜的土壤水分条件进行耦合,维持较高光合生产力的适宜水分条件为44.2%
2.3 不同水分条件下蒸腾速率光响应
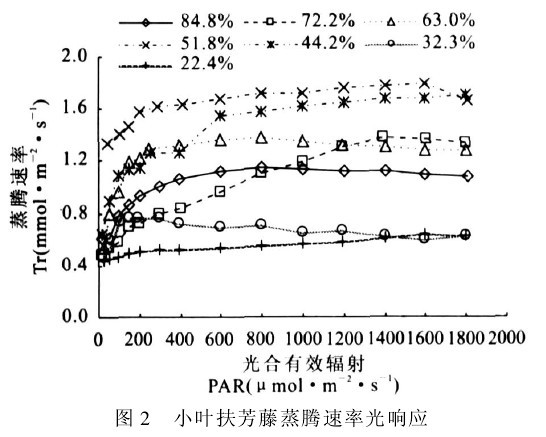
图2 小叶扶芳藤蒸腾速率光响应
图2表明,在各土壤水分条件下,蒸腾速率的光响应曲线表现为:低光照强度下(PAR<200μmol!m-2!s-1),随着PAR的增强,Tr上升较快:此后随着PAR的增加Tr缓慢上升。相同土壤湿度下,Tr对PAR变化的响应不太敏感,较强的PAR并不导致Tr的明显增加.但不同土壤湿度下的Tr值差别较大,在RWC为51.8%左右时,Tr值维持在最高水平:其中维持较高蒸腾速率的RWC在44.2%~72.2%之间,超出此土壤湿度范围后,无论RWC增加或降低,Tr值反而下降.Tr随RWC和PAR的变化规律表明,小叶扶芳藤蒸腾作用主要受土壤水分影响,受光照强度变化的影响较小.
2.4 不同水分条件下水分利用效率光响应
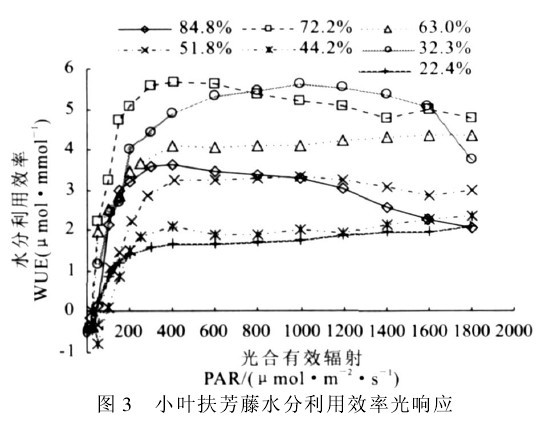
图3 小叶扶芳藤水分利用效率光响应
图3表明,各土壤墒情条件下,水分利用效率的光响应变化趋势类似于光合速率的光响应变化。低光强(PAR<400μmol!m-2!s-1)下,随着PAR的增强,WUE反应敏感,上升较快,随后WUE缓慢上升达到其光饱和点(在600~1 400μmol!m-2!s-1左右).此后WUE随着光强的增强变化较小,仅在高水分高光强条件下,下降幅度较大,表明小叶扶芳藤对光照强度的适应范围较广(在PAR为600~1 600μmol!m-2!s-1之间基本相同).但不同土壤湿度下的WUE差别明显,随着土壤湿度得增加,WUE有增加趋势,且在RWC为72.2%时WUE达到最大值,表明适度的水分下降在一定程度上利于水分利用效率的提高.在维持较高WUE的PAR(600~1 600μmol!m-2!s-1)范围内,WUE均值最大为5.67μmol!m-2!s-1(RWC为72.2%),其中MWC在51.8%~72.2%范围内,WUE维持在较高值3.25~5.67μmol!m-2!s-1,与维持WUE最低点(RWC为22. 4%)相比,增幅在54.8%~170.3%,而在低水分点(RWC为32.3%),WUE仍维持较高值.可见在土壤湿度偏高(RWC为Fig.3 Light responses of WUE ofE.fortuneivar.radicans84.8%)和偏低(RWC为32.3%)时,WUE都具有较高水平,表明小叶扶芳藤对土壤湿度的适应范围较广,维持高WUE的适宜RWC为32.3%~84.8%。
2.5 不同水分条件下光能利用效率光响应
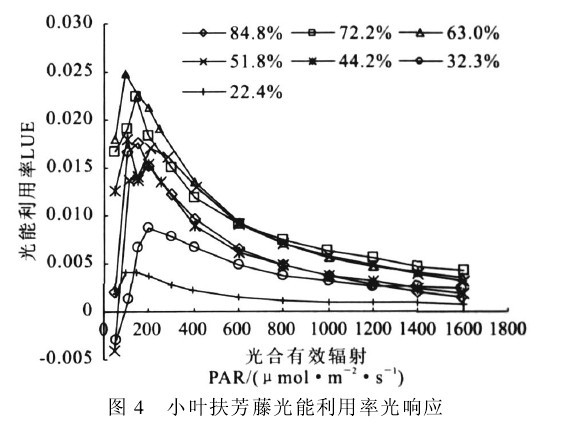
图4 小叶扶芳藤光能利用率光响应
图4表明,各土壤水分条件下,LUE的光响应过程基本相似,为明显的单峰曲线,变化幅度为0.004~0.025.低光强(PAR<200μmol!m-2!s-1)下,随着PAR的增强,LUE急剧上升,对光强反应敏感,随后在100~300μmol!m-2!s-1光强范围内,达到峰值.此后LUE逐渐下降,但在高光强下(PAR>800μmol!m-2!s-1)变化较小.土壤水分对LUE影响较大,在22.4%
2.6 不同土壤墒情条件下光合参数类型间差异
依据Pn、Tr、WUE、LUE 4个光合生理指标,对7个土壤湿度点进行聚类分析,当聚类数为3时,结果见表2.Ⅰ类含4个水分点,RWC在51.8%~72.2%和32.3%,为高光合速率、高蒸腾速率、较高水分利用效率、高光能利用效率型:Ⅱ类含1个水分点,RWC在44.2%左右,为较高光合速率、较高蒸腾速率、高水分利用效率、较高光能利用效率型:Ⅲ类含2个水分点,为低光合速率、低蒸腾速率、低水分利用效率、低光能利用效率型.方差分析表明,这3个水分点的Pn、Tr、WUE、LUE均值差异都极显著,说明土壤水分条件的不同,对小叶扶芳藤的苗期生理特性影响较大.通过聚类分析发现,土壤水分对Pn、Tr及LUE的影响具有很大的同步性,都表现为Ⅰ类水分群数值最高,Ⅱ类次之,Ⅲ类最低,仅WUE在Ⅱ类水分群最高,Ⅰ类次之,Ⅲ类最低.
3 讨 论
光合作用与蒸腾作用是植物对环境变化很敏感的生理过程,植物叶片对环境的光合响应提供了植物在不同光条件下生存和生长的能力以及对不断变化的环境条件适应能力的信息,它们既受外界因子的影响,也受植物体内部结构和生理状况的调节。水分因子是植物生长的最大限制因素,是光合作用的一种不可缺少的原料,同时,水分通过气孔蒸腾是蒸腾的主要方式,植物进行光合作用时作为原料消耗的水,只是植物从土壤中吸收的水量的很小一部分,其余的绝大部分都是通过蒸腾作用散失掉的.因此,水分亏缺时光合作用的降低,并不是由于水原料供应不足,而是由于水分亏缺引起的气孔或非气孔因素的限制。
本研究结果表明,小叶扶芳藤的光合作用、蒸腾作用及水分利用效率与土壤墒情和光照强度密切相关,而且有明显的阈值.从利于小叶扶芳藤进行高光合生产力和有效蒸腾的角度来确定WUE和LUE的适宜土壤水分和光照条件,RWC<32.3%时,苗木处于重度水分胁迫,Pn值较小:当土壤水分供应充足(RWC>72.2%)时,Pn不再随着土壤水分条件的改善而上升,甚至表现出一定的下降趋势,结合维持高LUE的土壤水分和光照范围,因此可以认为在44.2%
植物光合作用光饱和点与补偿点显示了植物叶片对强光和弱光的利用能力,代表了植物的需光特性和需光量.光补偿点较低、光饱和点较高的植物对光环境的适应性较强,反之适应性较弱。本研究表明,在适宜的土壤湿度范围内,小叶扶芳藤的LCP和Φ分别在22.6~28.0μmol!m-2!s-1之间和0.024~0.031 mol!mol-1左右,其LCP与典型阳性植物LCP(20~40μmol!m-2!s-1)相差不大:Φ值接近生长良好作物的Φ值(0.04~0.07 mol!mol-1之间)。饱和光强在PAR为1 000~1 400μmol/m-2s-1之间,并且在较宽的光照强度范围(PAR在600~1 600μmol!m-2!s-1)内,都具有较高的Pn和WUE水平.这表明小叶扶芳藤既有一定的耐荫性,又能适应较强的阳光辐射,是对光适应的生态幅较宽的植物。
植物水分利用效率作为评价植物对环境适应的综合生理生态指标,也是确定植物体生长发育所需水分供应的重要指标之一。当小叶扶芳藤遭受不利于光合作用的渍水或缺水逆境胁迫(RWC大于72.2%或小于44.2%)后,虽然光合速率下降,但蒸腾速率的下降幅度更大,因而即使在较高光强下仍能保持较高的水分利用效率.表明小叶扶芳藤具有高效利用水分的生理特征,在遭遇水分逆境后,能够通过自身生理调节(如大幅度减少蒸腾耗水等)来适应外界水分和光照条件的变化,延缓或减轻光合生理机构的损坏程度,维持一定的光合作用和较高的水分利用效率。
植物的干物质有90%来自光合作用,光能利用效率的大小是决定植物生产力高低的重要因素。本研究结果表明,随着土壤RWC(32.3%~72.2%)的递增,LCP降低,LSP和Φ升高,在RWC为72.2%时,达到LCP最低(22.6μmol!m-2!s-1),LSP最高(1 400μmol!m-2!s-1),说明在此水分条件下,小叶扶芳藤利用弱光和强光能力较强,有利于有机物质的积累,能适应多种光照环境.但土壤水分亏缺明显影响小叶扶芳藤叶片光合作用的LSP和Φ,因而在相同光量子密度情况下降低其叶片光能利用效率.可见合适的土壤水分能够提高小叶扶芳藤的光能利用效率和光照生态幅.因此在种植小叶扶芳藤时,为提高光能利用效率,可供应适宜的水分,同时可搭建爬棚,改善空间配置结构,能显著提高小叶扶芳藤生物产量和生存竞争力。
综上所述,小叶扶芳藤随着土壤墒情和光照强度的不同,有规律地调节自身的生理活动,以实现对水分和光照资源的充分利用.其光合生理参数的光响应值表现出较高的可塑性,说明其对光强和水分的适应能力较强,对逆境具有较高的潜在适应能力,这对小叶扶芳藤在干、湿环境及异质光环境中的生存和生长至关重要。